Eastern Mojave Vegetation
Eastern Mojave Vegetation
Tom Schweich
|
Eastern Mojave Vegetation
| Summaries of Literature About the Mid Hills. |
|
Tom Schweich |

Topics in this Article: Ecology of Three Yucca Species Patterns of Canopy Development and Carbon Gain Lei and Walkers's 1997 Papers Spacing Patterns in Mojave Desert Plant Communitie Joshua Tree Woodland North of Cima Study of Single-Needle Pinyons Vegetation of Caruthers Canyon Rocky Mountain White Fir (Abies concolor) Literature Cited |
(No Preface) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Other articles:
|
Ecology of Three Yucca Species | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Literature Cited:
Locations: Lobo Point. |
Yeaton et al. (1985) studied the three species association of Joshua Tree (Yucca brevifolia), Mohave Yucca (Y. schidigera), and Blue Yucca (Y. baccata) occupying a 700 m elevational gradient along Cima-Ivanpah Road (= Morning Star Mine Road) and Cedar Canyon Road, 13 to 36 km north of Lobo Point. In the course of their study, Yeaton et al. (1985) accomplished six tasks: | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1. Yucca Density Along An Elevational Gradient | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Literature Cited:
|
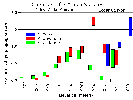
Yeaton et al. (1985) found that vegetation increases in density and complexity from lower elevations to higher. Species composition also changes constantly with increasing elevation from creosote bush scrub to pinyon-juniper woodland. This indicates that temperature decreases and rainfall increases with increasing elevation over this transect. Y. schidigera and Y. brevifolia are found together in the lower portion of the elevational gradient. Y. baccata replaces Y. schidigera at higher elevations. The maximum density and biomass for all three Yucca species totaled occurs at 1375 m. Y. schidigera and Y. brevifolia, which are associated extensively over the gradient, reach peaks of abundance toward the upper edges of their range but Y. baccata shows no change in its density or biomass from the upper portions of its range down to 1450 m where it drops out suddenly, to be replaced down slope by Y. schidigera. My chart which summarizes Yeaton et al. (1985) data is shown below: | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Locations: Cedar Canyon. |
A discontinuity occurs in the data when the transect enters Cedar Canyon, because factors other than elevation may cause changes in species density. Throughout Cedar Canyon a patchwork of Joshua tree woodland, pinyon-juniper woodland, and sagebrush scrub is observed. I believe this results from geomorphic expression of underlying geologic structure, and soil chemistry from three different rock types in the canyon. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2. Measured water potential for adjacent individuals at 1375 m. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Water potential measurements show that Y. schidigera and Y. baccata have water potentials of -20 to -22 bars and are more similar to each other than they are to Y. brevifolia with a measured water potential of -27 bars. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3. Measured transpiration rates. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Y. brevifolia has a significantly higher transpiration rate than Y. schidigera and Y. baccata which have very similar transpiration rates. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4. Measured temperature of upper leaf surfaces. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Upper leaf temperatures are generally lower for Y. brevifolia than for the other two species. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5. Counted density of stomata. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Y. brevifolia had fewer stomata per square millimeter than either Y. schidigera or Y. baccata. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6. Measured competitive interaction at the 1450 m site. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| "All intraspecific and interspecific pairings show significant positive, linear relationships between sums of sizes and distances apart of nearest neighbors," i.e., each species competes with other individuals of its species and with other species. Competition is greatest between Y. schidigera and Y. baccata. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Summary | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Literature Cited:
|
Y. schidigera and Y. baccata are so similar in appearance that for many years Y. schidigera was considered a tree-like form of Y. baccata. Physiological data reported here supports the notion that the two species are also very similar ecologically. Yeaton et al. (1985) proposed that the similarity of Y. schidigera and Y. baccata causes interspecific competition that limits the lower edge of the distribution of Y. baccata. They further proposed that the mode of competition between the two species is water utilization, in particular which subsurface zones might be used by the various species. Cody (1986a, 1986b), reviewed below, also discusses interspecific competition for water. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Other articles:
|
Patterns of Canopy Development and Carbon Gain | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Literature Cited:
Locations: Lobo Point. |
Comstock, Cooper and Ehleringer (1988) studied seasonal patterns of canopy development and carbon gain in nineteen warm desert shrub species. This study was initiated to collect baseline data on the phenology of carbon gain in warm desert species. A special emphasis was placed on contrasting the performance of photosynthetic leaves and twigs. The 19 species studied could be subdivided into six useful groups based on 1) the percent contribution of green twigs to whole plant carbon gain, and 2) the percent reduction of photosynthetic area during drought periods. The groupings, as I interpret them from the article text, are shown in Table 1. The species names are shown only for those species that also occur at Lobo Point. In those species that maintained a substantial canopy area through the drought period, previously stressed tissues showed substantial recovery after fall rains. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Table 1. Groupings of Plant Species by Photosynthetic Strategy.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Other articles:
|
Lei and Walkers's 1997 Papers | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Literature Cited:
|
Lei and Walker (1997a), Lei and Walker (1997b), and Lei (1997) published a series of related papers which appear to have been drawn from Lei's thesis research describing ecological aspects of Coleogyne communities in southern Nevada. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Classification and Ordination of Coleogyne Communities | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Literature Cited:
Locations: Las Vegas. Sheep Range. Spring Mountains. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Twelve elevational transects were located on the Spring Mountains and three were located on the Sheep Range. Each transect included 2 plots in the Larrea-Ambrosia shrubland just below the Coleogyne zone, extended through the entire Coleogyne zone and included 2 plots in the Pinus-Juniperus woodland just above the Coleogyne zone. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
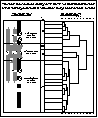 Interpretive diagram of Lei and Walkers results. Interpretive diagram of Lei and Walkers results.
|
TWINSPAN (2-way indicator species analysis) identified 4 primary species and stand groups from the 2 mountain ranges (extensive survey). They were: Larrea-Ambrosia, Coleogyne, upper Coleogyne, and Pinus-Juniperus. Yucca baccata and Y. brevifolia were common in the Coleogyne zone, along with Prunus fasciculata and Thamnosma montana. Gutierrezia sarothrae was particularly abundant in the upper Coleogyne zone which also contained Atriplex canescens and Chrysothamnus nauseosus. My interpretation of Lei and Walker's (1997a) transect plan and their species dendrogram is shown at right. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Analysis of DECORANA (detrended correspondence analysis) results indicated that elevation and soil depth were the environmental factors most significantly associated with distribution of species and stand groups. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Locations: Lucky Strike Canyon. Spring Mountains. |
In addition, the Lower Coleogyne elevational boundary was analyzed in detail in Lucky Stike Canyon, on the western edge of the Spring Mountains with six replicate transects including one plot just below the lower elevational limit of Coleogyne and five plots each at increasing elevations in the Coleogyne zone. The Coleogyne zone was subdivided into pure stands and upper and lower ecotones for further investigation of species distribution and environmental factors. Elevation was the only significant physical factor associated with distribution of species and stand groups in the detailed survey in Lucky Strike Canyon. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Biotic and Abiotic Factors Influencing the Distribution of Coleogyne Communities | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Literature Cited:
Locations: Las Vegas. Lucky Strike Canyon. |
In their second paper, Lei and Walker (1997b) characterized the extent that environmental factors correlate with Coleogyne density and examined variation in biotic and abiotic factors along the lower elevational boundary of Coleogyne in Lucky Strike Canyon near Las Vegas, Nevada. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A complete review of their results is beyond the scope of this summary. However, they found that mean density of Coleogyne increased significantly with elevation. Coleogyne density was positively correlated with gravimetric soil moisture, soil organic matter, Coleogyne water potential, Coleogyne stem and leaf phosphorus, and Coleogyne leaf biomass. However, Coleogyne density was negatively correlated with soil temperatures, soil compaction, and Coleogyne stem and leaf nitrogen. Coleogyne stem biomass and elongation were generally negatively correlated with Coleogyne density. Coleogyne density was weakly correlated with soil pH, soil depth, soil nitrogen, soil and leaf phosphorus; these variables did not exhibit a consistent pattern with increasing elevation. In summary, edaphic factors, particularly, soil moisture and soil organic matter appear to play a major role in determining the distribution of Coleogyne shrublands in southern Nevada. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Variation in Germination Response | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Literature Cited:
|
Lei (1997) tested germination of approximately 9600 Blackbrush seed from 8 locations and two elevations (120 m and 1550 m) in the Mojave Desert. The seeds were treated with various combinations of prechill, incubation temperature and watering frequency. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Germination of blackbrush seeds was maximal when preceded by a prechill period of 4-6 weeks. Seeds incubated at room temperature germinated poorly. Seeds collected at warm, low elevation sites appeared to be less dormant (required less prechill time), germinated faster, and showed a higher overall germination response at low temperature relative to cold, high elevation sites. Frequencies of watering also determined the germination response; watering at 2 week intervals resulted in the greatest germination. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Synonymy | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Forsellesia nevadensis = Glossopetalon spinescens
|
|
Other articles:
| Spacing Patterns in Mojave Desert Plant Communitie
|
Literature Cited:
Locations: Granite Mountains. Mid Hills.
Cody (1986a) studied shrub spacing patterns at the Granite Mountains and in the Mid Hills. Five species show uniform distributions, 30 species show a clumped distribution, and 32 species are distributed randomly. Top ranking species in volume tend toward uniform spacing. The larger species trend toward more uniform spacing at sites where they reach higher densities, see Table 2. Some smaller species tend toward increasing aggregation as species density increases. Other smaller species are randomly scattered when rare and clumped when common.
|
|
Literature Cited:
Table 2. Representative conclusions from Cody (1986a).
|
|
Literature Cited:
Cody's (1986a) data also shows that some species have certain other species as neighbors far more often than would be expected by chance, and others occur less often than expected by chance. Cody discusses two possible causes: germination opportunity, and root interaction. Cody (1986b) reports direct observations of desert shrub root system morphology. A great deal of interspecific variability exists, along with a great deal of intraspecific constancy. He concludes that space preemption occurs
among species that compete for a single limiting resource, such as water. Shrub species interact over space and presumably over the water rights included within this space. Therefore, a substantial component of the shrub diversity in this vegetation may be owed to a diversity of water uptake strategies, from strictly soil surface root systems, e.g.,Hedgehog Cactus (Echinocereus engelmannii) to strictly deep tap-rooted system, e.g., Burrobrush (Hymenoclea salsola), and including all kinds of intermediate morphologies.
|
|
Other articles:
| Joshua Tree Woodland North of Cima
|
Literature Cited:
As reported in Vasek and Barbour (1988), Vasek and Rowlands (not published)
measured vegetational characteristics of a Joshua tree woodland north of Cima. This
would be on the south slope of Cima Dome. Elevation is not reported. However, the
elevation of Cima is 1273 m and the top of Cima Dome is 1751 m. The dominant
species by far is Ericameria [Happlopappus] cooperi. Other common species are
Winter Fat (Krascheninnikovia [Eurotia] lanata), Ephedra nevadensis, and Bladder
Sage (Salazaria mexicana). Joshua Tree (Yucca brevifolia) is visually dominant,
resulting in the vegetation type being called Joshua tree woodland, but the species
scores low on quantitative measures of dominance.
|
|
Literature Cited:
It is not usual to re-publish someone else's data in entirety. However, the data in this article is very much relevant to my current study. It is one of the few published studies of eastern Mojave vegetation. It also is the only measured plot of Joshua tree woodland referenced in Sawyer and Keeler-Wolf (1995) and the data in this table will be needed to analyze regional trends and definitions of vegetation associations and series.
|
|
| --------------------------------------------------------------- Taxon Density Cover(%) Frequency IV --------------------------------------------------------------- Escobaria vivipara 1 t 1 0.59 Echinocereus engelmannii 5 0.01 5 5.05 Ephedra nevadensis 18 0.49 14 12.23 Eurotia lanata 23 0.43 13 11.99 Ericameria cooperi 164 1.61 38 49.06 Ericameria linearifolius 6 0.12 4 3.42 Hilaria rigida 488 2.72 40 96.92 Hymenoclea salsola 12 0.86 11 12.69 Lycium andersonii 5 0.86 5 6.94 Lycium cooperi 9 0.48 8 8.22 Menodora spinescens 12 0.68 8 10.00 Mirabilis bigelovii 1 0.01 1 0.64 [= M. laevis var. villosa] TAS 19-Feb-05 Muhlenbergia porteri 28 0.10 18 12.57 Opuntia acanthocarpa 1 t 1 0.59 Opuntia echinocarpa 1 t 1 0.59 Opuntia ramosissima 9 1.08 8 12.48 Salazaria mexicana 21 2.33 15 26.15 Salvia dorrii 2 0.07 1 1.21 Stipa speciosa 6 0.01 5 3.11 Thamnosma montana 10 0.68 8 9.80 Yucca baccata 3 1.64 3 13.38 Yucca brevifolia 5 0.20 5 4.36 Total 830 14.08 ---------------------------------------------------------------
|
Other articles:
| Study of Single-Needle Pinyons
|
Literature Cited:
Locations: Granite Mountains. Great Basin. New York Mountains. Providence Mountains. Wild Horse Mesa.
Bailey (1988) reported on his extensive investigation of pinyons with single-needle fascicles whose natural ranges lie mainly in the Californias and the Great Basin. Characters used by Bailey are number of resin ducts, number of stomatal lines, fascicle sheath curl back, and endosperm composition. He recognized three taxa with limited geographical overlap. Bailey's Collection No. 241 was made at the north end of Wild Horse Mesa. His sample showed no individuals of P. monophylla, and four individuals each of P. californiarum and P. californiarum subsp. fallax. Along the axis of the G ranite Mountains, Providence Mountains and New York Mountains, few individuals of P. monophylla are found. Most common is P. californiarum, with P. c. subsp. fallax in smaller numbers and found mainly in the central and northern sections of the mountain
range axis.
|
|
Other articles:
| Vegetation of Caruthers Canyon
|
Literature Cited:
Locations:
Caruthers Canyon.
Clark Mountain.
Lobo Point.
New York Mountains.
In another study about a close-by area, F. C. Vasek and H. B. Johnson (not published, reported in
Vasek and Thorne, 1988) measured the relative densities of vegetation at sites in Caruthers Canyon, New York Mountains and on Clark Mountain. Caruthers Canyon ranges from 1650 to 2100 in altitude and is 22 northeast of Lobo Point. Vasek and Johnson's data showed presence of Coffeeberry (Rhamnus californica), Redberry (R. crocea), Desert Ceanothus (Ceanothus greggii vestitus), Yerba Santa (Eriodictyon augustifolia) and Desert Olive (Forestiera neomexicana). The vegetation found in Caruthers Canyon suggests that it is unique among reported eastern Mojave sites. This is the only other study that reported Wright's Buckwheat (Eriogonum wrightii) found among the pinyons at Lobo Point. Perhaps the R. crocea reported by Vasek and Johnson was R. crocea ssp. ilicifolia, now renamed R. ilicifolia and found at North Wild Horse Mesa.
|
|
Other articles:
| Rocky Mountain White Fir (Abies concolor)
|
Locations: Clark Mountain. Kingston Range. New York Mountains. Spring Mountains.
At even higher elevations, the Rocky Mountain White Fir
(Abies concolor var. concolor)
grove in the New York Mountains has achieved legendary status.
It contains about 30 trees in 0.8 ha at an elevation of 2100 m
(Hendrickson and Prigge, 1975).
White fir is also found in the Kingston Mountains and Clark Range
of California and the Charleston Mountains (=Spring Mountains) of Nevada.
|
|
| Literature CitedA list of all literature cited by this web site can be found in the Bibliography. |
Bailey, D. K. 1988.
The Single-needle Pinyons - One Taxon or Three?.
pp. pp. 1-18. in Hall, Clarence A., Jr and Victoria Doyle-Jones (Eds.). 1988..
The Mary DeDecker Symposium. University of California, White Mountain
Research Station..
Plant Biology of Eastern California.
| |
Cody, M. L. 1986a.
Spacing patterns in Mojave Desert plant communities: near-neighbor analysis.
Journal of Arid Environments.
11: 199-217.
{TAS}
| |
Cody, M. L. 1986b.
Structural niches in plant communities.
pp. 381-405 in Diamond, J., and T. J. Case (Eds.).
Community Ecology.
San Francisco: Harper & Row.
{TAS}
| |
Comstock, J. P., T. A. Cooper, and J. R. Ehleringer. 1988.
Seasonal patterns of canopy development and carbon gain
in nineteen warm desert shrub species.
Oecologia (Berlin).
75(3):327-335.
{TAS}
[Ecophysiology Lab,
Department of Biology,
University of Utah, Salt Lake City, UT 84112 USA]
| |
Hickman, James C. (Ed.). 1993.
The Jepson manual: higher plants of California.
Berkeley, CA: University of California Press..
| |
Lei, Simon A. and Lawrence R. Walker. 1997a.
Classification and ordination of Coleogyne communities in southern Nevada.
Great Basin Naturalist.
57(2):155-162.
| |
Lei, Simon A., and Lawrence R. Walker. 1997b.
Biotic and abiotic factors influencing the distribution of Coleogyne communities in southern Nevada.
Great Basin Naturalist.
57(2):163-171.
| |
Lei, Simon A. 1977.
Variation in germination response to temperature and water availability in blackbrush (Coleogyne ramosissima) and its ecological significance.
Great Basin Naturalist.
57(2):172-177.
| |
Sawyer, John O., and Todd Keeler-Wolf. 1995.
A Manual of California Vegetation.
Sacramento, CA: California Native Plant Society.
471 p.
| |
Vasek, F. C., and M. G. Barbour. 1988.
Mojave Desert scrub vegetation.
pp. pp. 835-868 in Barbour, Michael G., and Jack Major. 1988.
Terrestrial Vegetation of California, New Expanded Edition.
Special Publication Number 9..
1988.
{TAS}
| |
Vasek, F. C., and R. F. Thorne. 1988.
Transmontane coniferous vegetation.
pp. pp. 797-832 in Barbour, Michael G., and Jack Major. 1988.
Terrestrial Vegetation of California, New Expanded Edition.
Special Publication Number 9.
1988.
| |
Yeaton, R. I., R. W. Yeaton, J. P. Waggoner III and J. E. Horenstein. 1985.
The ecology of Yucca (Agavaceae) over an environmental gradient in the Mohave Desert: distribution and interspecific interactions.
Journal of Arid Environments.
8: 33-44.
|
If you have a question or a comment you may write to me at:
tomas@schweich.com
I sometimes post interesting questions in my FAQ, but I never disclose your full name or address.
|
|
Date and time this article was prepared: 5/12/2025 2:56:50 PM |